
- Положення вірусів в системі живого
- походження вірусів
- структура
- механізм інфікування
- Класифікація
- ICTV класифікація
- Класифікація Балтімора
- Історія
- застосування вірусів
- Примітки
?
віруси 
ротавірус
наукова класифікація Латинська назва Virae Позасистемні групи
по Балтімор
I. дцДНК-містять
II. оцДНК-містять
III. дцРНК-містять
IV. (+) ОцРНК-містять
V. (-) оцРНК-містять
VI. оцРНК-містять ретровіруси
VII. дцДНК-містять ретроідние віруси
Вірус (від лат. virus - отрута) - мікроскопічна частка, здатна інфікувати клітини живих організмів. Віруси є облігатними паразитами - вони не здатні розмножуватися поза клітиною. В даний час відомі віруси, що розмножуються в клітинах рослин , тварин , грибів і бактерій (Останніх зазвичай називають бактеріофагами ). Виявлено також вірус, що вражає інші віруси [1] . Віруси є молекули нуклеїнових кислот ( ДНК або РНК ), Ув'язнені в захисну білкову оболонку ( капсид ). Наявність капсида відрізняє віруси від інших інфекційних агентів, віроідов . Віруси містять тільки один тип нуклеїнової кислоти: або ДНК , або РНК . Раніше до вірусів також помилково відносили пріони , Проте згодом виявилося, що ці збудники є особливими білки і не містять нуклеїнових кислот.
Положення вірусів в системі живого
Віруси є однією з найпоширеніших форм існування органічної матерії на планеті за чисельністю: води світового океану містять колосальну кількість бактеріофагів (около 1011 частинок на мілілітр води).
Віруси мають генетичні зв'язки з представниками флори і фауни Землі. Згідно з останніми дослідженнями, геном людини більш ніж на 32% складається з інформації, кодируемой вірус-подібними елементами і транспозонами . За допомогою вірусів може відбуватися так званий горизонтальний перенос генів ( ксенологія ), Тобто передача генетичної інформації не від батька до сина і так далі, а між двома несумісними (або навіть відносяться до різних видів) особинами. Так в геномі вищих приматів існує білок сінцітін, який, як вважається, був привнесений ретровирусом . Іноді віруси утворюють з тваринами симбіоз [2] . Так, наприклад, отрута деяких паразитичних ос містить структури, звані полі-ДНК-вірусами (Polydnavirus, PDV), що мають вірусне походження.
походження вірусів
Віруси - збірна група, яка не має загального предка. В даний час існує декілька гіпотез, що пояснюють походження вірусів.
Вважається, що великі ядерно-цитоплазматичні ДНК-віруси походять від більш складних (і, можливо, клітинних, таких як сучасні мікоплазми та рикетсії ), Внутрішньоклітинних паразитів, які втратили значну частину свого геному. І дійсно, деякі великі ДНК-віруси ( мімівіруса , вірус віспи ) Кодують функціонально надлишкові на перший погляд ферменти, мабуть, залишилися їм у спадок від більш складних форм існування. Слід також зазначити, що деякі вірусні білки не виявляють ніякої гомології з білками бактерій , архей і еукаріот , Що свідчить про порівняно давньому відокремленні цієї групи.
ДНК-містять бактеріофаги і деякі ДНК-віруси еукаріот, можливо походять від мобільних елементів, ділянок ДНК здатних до самостійного реплікації в клітині.
Походження деяких РНК-вірусів пов'язують з віроїди . Віроїди представляють собою високо структуровані кільцеві фрагменти РНК, реплицируемой клітинної РНК-полімераза . Вважається, що віроїди представляють собою «втекли інтрони »- вирізані в ході сплайсингу незначущі ділянки мРНК , Які випадково придбали здатність до реплікації. Білків віроїди не кодують. Вважається, що придбання віроїди кодують ділянок ( відкритої рамки зчитування ) І призвело до появи перших РНК-вірусів. І дійсно, відомі приклади вірусів, що містять виражені віроїди-подібні ділянки (вірус гепатит А Дельта).
структура
Вірусні частинки ( віріони ) Представляють собою білкову капсулу - капсид , Що містить геном вірусу, представлений однією або декількома молекулами ДНК або РНК . Капсид побудований з капсомеров - білкових комплексів, що складаються в свою чергу з протомеров. Нуклеїнова кислота в комплексі з білками позначається терміном нуклеокапсид. Деякі віруси мають також зовнішню ліпідну оболонку. Розміри різних вірусів коливаються від 20 ( пікорнавіруси ) До 500 ( мімівіруса ) и більше нанометрів . Віріони часто мають правильну геометричну форму ( ікосаедр , циліндр ). Така структура капсида передбачає ідентичність зв'язків між складовими її білками, і, отже, може бути побудована з стандартних білків одного або декількох видів, що дозволяє вірусу економити місце в геномі.
механізм інфікування
Умовно процес вірусного інфікування в масштабах однієї клітини можна розбити на кілька взаімоперекривающіхся етапів:
- Приєднання до клітинної мембрани - так звана адсорбція . Зазвичай для того, щоб віріон адсорбувався на поверхні клітини, вона повинна мати у складі своєї плазматичноїмембрани білок (часто глікопротеїн ) - рецептор, специфічний для даного вірусу. Наявність рецептора нерідко визначає коло господарів даного вірусу, а також його тканеспеціфічность .
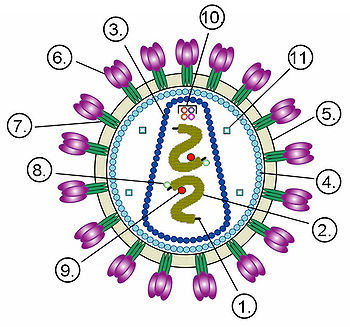
Структура віріона неікосаедріческого оболочечного вірусу на прикладі ВІЛ .
Цифрами позначені: (1) РНК-геном вірусу, (2) нуклеокапсид, (3) капсид, (4) білковий матрикс, що підстилає (5) липидную мембрану , (6) gp120 - глікопротеїн, за допомогою якого відбувається зв'язування вірусу з клітинною мембраною, (7) gp41 - трансмембранний глікопротеїн.
Цифрами 8-11 позначені білки, що входять до складу віріона і необхідні вірусу на ранніх стадіях інфекції: (8) - інтеграли, (9) - зворотна транскриптаза , (10) - Vif, Vpr, Nef і p7, (11) - протеаза .
- Проникнення в клітину. На наступному етапі вірусу необхідно доставити всередину клітини свою генетичну інформацію. Деякі віруси привносять також власні білки, необхідні для її реалізації (особливо це характерно для вірусів, що містять негативні РНК). Різні віруси для проникнення в клітину використовують різні стратегії: наприклад, пікорнавіруси впорскують свою РНК через плазматичну мембрану, а віріони ортомиксовирусов захоплюються клітиною в ході ендоцитозу , Потрапляють в кисле середовище лізосом , Де відбувається їх остаточне дозрівання (депротеїнізація вірусної частинки), після чого РНК в комплексі з вірусними білками долає лізосомальну мембрану і потрапляє в цитоплазму. Віруси також розрізняються за локалізацією їх реплікації, частину вірусів (наприклад, ті ж пікорнавіруси ) Розмножується в цитоплазмі клітини, а частина (наприклад, ортоміксовіруси) в її ядрі .
- Перепрограмування клітини. При зараженні вірусом в клітці активуються спеціальні механізми противірусного захисту. Заражені клітини починають синтезувати сигнальні молекули - інтерферони , Що переводять навколишні здорові клітини в противірусну стан і активують системи імунітету. Пошкодження, що викликаються розмноженням вірусу в клітці, можуть бути виявлені системами внутрішнього клітинного контролю, і така клітина повинна буде «покінчити життя самогубством» в ході процесу, званого апоптозом або програмованої клітинної смерті. Від здатності вірусу долати системи противірусного захисту безпосередньо залежить його виживання. Не дивно, що багато вірусів (наприклад, пікорнавіруси , Флавівіруси) в ході еволюції набули здатності пригнічувати синтез інтерферонів, апоптозну програму і так далі. Крім придушення противірусного захисту, віруси прагнуть створити в клітці максимально сприятливі умови для розвитку свого потомства. Хрестоматійним прикладом перепрограмування систем клітини-господаря є трансляція РНК ентеровірусів (сімейство пікорнавіруси ). Вірусна протеаза розщеплює клітинний білок eIF4G, необхідний для ініціації трансляції переважної більшості клітинних мРНК (що транслюються по так званому кеп-залежному механізму). При цьому ініціація трансляції РНК самого вірусу відбувається іншим способом ( IRES -залежний механізм), для якого цілком достатньо відрізаного фрагмента eIF4G. Таким чином, вірусні РНК набувають ексклюзивні «права» і не конкурують за рибосоми з клітинними.
- Персистенція. Деякі віруси можуть переходити в латентний стан (так звана персистенция для вірусів еукаріот або лізогенія для бактеріофагів - вірусів бактерій ), Слабо втручаючись в процеси, що відбуваються в клітині, і активуватися лише за певних умов. Так побудована, наприклад, стратегія розмноження деяких бактеріофагів - до тих пір поки заражена клітина знаходиться в сприятливому середовищі, фаг не вбиває її, успадковується дочірніми клітинами і нерідко інтегрується в клітинний геном. Однак при попаданні зараженої лізогенним фагом бактерії в несприятливе середовище, збудник захоплює контроль над клітинними процесами так, що клітина починає виробляти матеріали, з яких будуються нові фаги (так звана літична стадія). Клітка перетворюється на фабрику, здатну виробляти багато тисяч фагів. Зрілі частинки, виходячи з клітини, розривають клітинну мембрану , Тим самим вбиваючи клітину. З персистенцією вірусів (наприклад, паповавирусов) пов'язані деякі онкологічні захворювання .
- Створення нових вірусних компонентів. Розмноження вірусів в найзагальнішому випадку передбачає три процеси - 1) транскрипція вірусного генома - тобто синтез вірусної мРНК, 2) її трансляція, тобто синтез вірусних білків і 3) реплікація вірусного генома (в деяких випадках, коли генетична інформація вірусу закодована у вигляді РНК геномна РНК одночасно грає роль мРНК, і, отже, процес транскрипції в паразітіруемой клітці не відбувається за непотрібністю). У багатьох вірусів існують системи контролю, що забезпечують оптимальне витрачання біоматеріалів клітини-господаря. Наприклад, коли вірусної мРНК накопичено достатньо, транскрипція вірусного генома пригнічується, а реплікація навпаки - активується.
- Дозрівання віріонів і вихід з клітини. Зрештою, новосинтезовані геномні РНК або ДНК одягаються відповідними білками і виходять з клітини. Слід сказати, що активно розмножується вірус не завжди вбиває клітину-господаря. У деяких випадках (наприклад, ортоміксовіруси) дочірні віруси відгалужуються від плазматичної мембрани, не викликаючи її розриву. Таким чином, клітина може продовжувати жити і продукувати вірус.
Класифікація
В таксономії живої природи віруси виділяються в окремий таксон Vira, який утворює в класифікації Systema Naturae 2000 разом з доменами Bacteria, Archaea і Eukaryota кореневої таксон Biota. Протягом XX століття в систематиці висувалися пропозиції про створення виділеного таксона для неклітинних форм життя (Aphanobionta Novak, 1930; надцарство Acytota Jeffrey, 1971; Acellularia), проте такі пропозиції не були кодифіковані.
Систематику і таксономії вірусів кодифікує і підтримує Міжнародний Комітет по таксономії Вірусів (International Committee on Taxonomy of Viruses, ICTV), що підтримує також і таксономічну базу The Universal Virus Database ICTVdB.
ICTV класифікація
Міжнародним Комітетом по таксономії Вірусів в 1966 році була прийнята система класифікації вірусів заснована на відмінності типу (РНК і ДНК), кількості молекул нуклеотіческіх кислот (одно- і двох-цепочечние) і на наявність чи відсутність оболонки ядра. Система класифікації являє собою серію ієрархічних таксонів :
загін (-Virales) сімейство (-Viridae) підродина (-Virinae) рід (-Virus) вид (-Virus)
Класифікація Балтімора
Нобелівський лауреат, біолог Девід Балтімор, запропонував свою схему класифікації вірусів, грунтуючись на відмінностях в механізмі продукції мРНК .Ця система включає в себе сім основних груп: [3] [4]
- (I) Віруси, що містять двуцепочечную ДНК і не мають РНК-стадії (наприклад, герпесвіруси , Поксвирусов, паповавіруси, мімівіруса ).
- (II) Віруси, що містять двуцепочечную РНК (наприклад, ротавіруси).
- (III) Віруси, що містять одноланцюжкові молекулу ДНК (наприклад, парвовіруси).
- (IV) Віруси, що містять одноланцюжкові молекулу РНК позитивної полярності (наприклад, пікорнавіруси , Флавівіруси).
- (V) Віруси, що містять одноланцюжкові молекулу РНК негативної або подвійний полярності (наприклад, ортоміксовіруси, філовірусів ).
- (VI) Віруси, що містять одноланцюжкові молекулу РНК і мають в своєму життєвому циклі стадію синтезу ДНК на матриці РНК, ретровіруси (Наприклад, ВІЛ ).
- (VII) Віруси, що містять двуцепочечную ДНК і мають в своєму життєвому циклі стадію синтезу ДНК на матриці РНК, ретроідние віруси (наприклад, вірус гепатиту B ).
В даний час, для класифікації вірусів використовуються обидві системи одночасно, як доповнюють один одного. [5] [6] [7]
Подальше поділ проводиться на основі таких ознак як структура генома (наявність сегментів, кільцева або лінійна молекула), генетична схожість з іншими вірусами, наявність ліпідної оболонки, таксономічна приналежність організму-господаря і так далі.
Історія
В 2002 в університеті Нью-Йорка був створений перший синтетичний вірус ( вірус поліомієліту ).
застосування вірусів
Див. також
посилання
література
- Mayo MA, Pringle CR Virus taxonomy - +1997 // Journal of General Virology. - 1998. - № 79. - С. 649-657.
Примітки
ресурси
Науково-популярні роботи
Wikimedia Foundation. 2010 року.











