
слайд №2  Сучасне вивчення структури білка
Сучасне вивчення структури білка
У XXI столітті дослідження білків перейшло на якісно новий рівень, коли досліджуються не тільки індивідуальні білки, а й одночасна зміна кількості і пост трансляційні модифікації великого числа білків окремих клітин, тканин або організмів. Ця область біохімії називається протеомікою.
За допомогою методів біоінформатики стало можливо не тільки обробити дані рентгеном-структурного аналізу, але і передбачити структуру білка, грунтуючись на його амінокислотної послідовності.
В даний час електронна мікроскопія великих білкових комплексів і передбачення малих білків і доменів великих білків за допомогою комп'ютерних програм по точності наближаються до вирішення структур на атомному рівні.
слайд №3  Первинна структура білків
Первинна структура білків
Молекули білка тривимірні і мають кілька рівнів організації.
Первинна структура - порядок чергування (послідовність) амінокислот в поліпептидному ланцюзі, з'єднаних між собою пептидними зв'язками.
Первинна структура індивідуальна для різних білків.
слайд №4  Лінійні поліпептидні ланцюги білків за рахунок взаємодії функціональних груп амінокислот набувають певну просторову структуру, яка називається «конформація».
Лінійні поліпептидні ланцюги білків за рахунок взаємодії функціональних груп амінокислот набувають певну просторову структуру, яка називається «конформація».
Всі молекули білків, що мають однакову первинну структуру мають однакову конформацію.
У білках розрізняють 2 основних типи конформації поліпептидних ланцюгів: вторинну і третинну структури.
Конформація білків Слайд №5 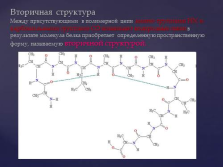 Вторинна структураМежду присутніми в полімерного ланцюга аміно-групами HN і карбонільних групами CO виникають водневі зв'язки в результаті молекула білка набуває певну просторову форму, звану вторинної структурою. слайд №6
Вторинна структураМежду присутніми в полімерного ланцюга аміно-групами HN і карбонільних групами CO виникають водневі зв'язки в результаті молекула білка набуває певну просторову форму, звану вторинної структурою. слайд №6  Механізм виникнення водневих зв'язків можна показати на прикладі взаємодії двох молекул води. У диполі води, як відомо, надлишок позитивних зарядів припадає на атоми водню, а надлишок негативних - на атоми кисню. При достатньому зближенні двох молекул води виникає електростатичне взаємодія між атомом кисню однієї молекули і атомом водню другий молекули води. Наслідком цього є ослаблення зв'язку між атомами водню і кисню в кожній молекулі води і відповідно виникнення нової, неміцною зв'язку (відзначена пунктиром) між атомом водню першої молекули і атомом кисню другий молекули води. Цю неміцну зв'язок прийнято позначати водневої зв'язком. слайд №7
Механізм виникнення водневих зв'язків можна показати на прикладі взаємодії двох молекул води. У диполі води, як відомо, надлишок позитивних зарядів припадає на атоми водню, а надлишок негативних - на атоми кисню. При достатньому зближенні двох молекул води виникає електростатичне взаємодія між атомом кисню однієї молекули і атомом водню другий молекули води. Наслідком цього є ослаблення зв'язку між атомами водню і кисню в кожній молекулі води і відповідно виникнення нової, неміцною зв'язку (відзначена пунктиром) між атомом водню першої молекули і атомом кисню другий молекули води. Цю неміцну зв'язок прийнято позначати водневої зв'язком. слайд №7  Найбільш поширені два типи вторинної структури білків. Перший варіант, званий a-спіраллю, реалізується за допомогою водневих зв'язків всередині однієї полімерної молекули. Водневі зв'язки формуються між 1-й і 4-й амінокислотами.
Найбільш поширені два типи вторинної структури білків. Перший варіант, званий a-спіраллю, реалізується за допомогою водневих зв'язків всередині однієї полімерної молекули. Водневі зв'язки формуються між 1-й і 4-й амінокислотами.
В результаті стягивающего дії водневих зв'язків молекула набуває форму спіралі - так звана a-спіраль, її зображують у вигляді зігнутої спиралевидной стрічки, що проходить через атоми, що утворюють полімерну ланцюг.
a-спіраль являє собою найжорсткіший тип вторинної структури, переважає в багатьох білках. слайд №8 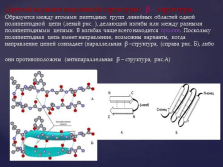 Інший варіант вторинної структури -? - структура. Утворюється між атомами пептидних груп лінійних областей однієї поліпептидного ланцюга (лівий рис.), Що робить вигини або між різними поліпептидними ланцюгами. У вигинах найчастіше знаходиться пролин. Оскільки поліпептидний ланцюг має напрямок, можливі варіанти, коли напрямок ланцюгів збігається (паралельна? -Структура, (праворуч рис. Б), або вони протилежні (антипаралельними? - структура, ріс.А) Слайд №9
Інший варіант вторинної структури -? - структура. Утворюється між атомами пептидних груп лінійних областей однієї поліпептидного ланцюга (лівий рис.), Що робить вигини або між різними поліпептидними ланцюгами. У вигинах найчастіше знаходиться пролин. Оскільки поліпептидний ланцюг має напрямок, можливі варіанти, коли напрямок ланцюгів збігається (паралельна? -Структура, (праворуч рис. Б), або вони протилежні (антипаралельними? - структура, ріс.А) Слайд №9  ? -Структура (по Березову Т.Т.) - складчастий тип, водневі зв'язки формують гофровану структуру з поліпептидного ланцюга. На схемах зображується у вигляді стрілки від N до С - кінця. слайд №10
? -Структура (по Березову Т.Т.) - складчастий тип, водневі зв'язки формують гофровану структуру з поліпептидного ланцюга. На схемах зображується у вигляді стрілки від N до С - кінця. слайд №10  У білках відзначають області з нерегулярною структурою білка, які часто називають «безладні клубки» Зміст різних типів вторинних структур
У білках відзначають області з нерегулярною структурою білка, які часто називають «безладні клубки» Зміст різних типів вторинних структур
1. Містять тільки a- спіралі
(Hb і міоглобін)
2. Містять a-спіралі і? - структури. (Лактатдегидрогеназа)
3. Містять тільки? - структури.
4. Мало регулярних вторинних структур.
слайд №11  Супервторічние структури.
Супервторічние структури.
Специфічний порядок формування вторинних структур називають супервторічной структурою білка.
розрізняють:
тип a /? - бочки, «цинковий палець», лейцінових застібка і ін. Слайд №12 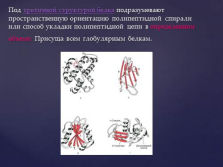 Під третинної структурою білка розуміють просторову орієнтацію поліпептидного спіралі або спосіб укладання поліпептидного ланцюга в певному обсязі. Властива всім глобулярним білків. слайд №13
Під третинної структурою білка розуміють просторову орієнтацію поліпептидного спіралі або спосіб укладання поліпептидного ланцюга в певному обсязі. Властива всім глобулярним білків. слайд №13  Типи нековалентних зв'язків, що стабілізують третинну структуру белка.1 - іонні зв'язку; 2 - воднева зв'язок; 3 - гідрофобні взаємодії неполярних груп; 4 - дисульфідний (ковалентний) зв'язок. слайд №14
Типи нековалентних зв'язків, що стабілізують третинну структуру белка.1 - іонні зв'язку; 2 - воднева зв'язок; 3 - гідрофобні взаємодії неполярних груп; 4 - дисульфідний (ковалентний) зв'язок. слайд №14  Найбільш важливі водневі зв'язки утворюються між атомом водню, що несе частковий позитивний заряд, і негативно зарядженим атомом кисню в білкової молекулі. Нижче представлені приклади водневих зв'язків, які можуть ще утворюватися в білкової молекулі: а) між пептидними ланцюгами; б) між двома гідроксильних груп; в) між іонізованої СООН-групою і ОН-групою тирозину; г) між ОН-групою серину і пептидного зв'язком. слайд №15
Найбільш важливі водневі зв'язки утворюються між атомом водню, що несе частковий позитивний заряд, і негативно зарядженим атомом кисню в білкової молекулі. Нижче представлені приклади водневих зв'язків, які можуть ще утворюватися в білкової молекулі: а) між пептидними ланцюгами; б) між двома гідроксильних груп; в) між іонізованої СООН-групою і ОН-групою тирозину; г) між ОН-групою серину і пептидного зв'язком. слайд №15  Дисульфідні зв'язки в структурі інсуліну людини Слайд №16
Дисульфідні зв'язки в структурі інсуліну людини Слайд №16  Першим білком, третинна структура якого була з'ясована Дж. Кендра на підставі рентгеноструктурного аналізу, виявився миоглобин кашалота. Це порівняно невеликий білок з мовляв. м. 16700, що містить 153 амінокислотних залишку (повністю з'ясована первинна структура), представлений однією поліпептидного ланцюгом. Основна функція міоглобіну - перенесення кисню в м'язах. Поліпептидний ланцюг міо-глобіну представлена у вигляді зігнутої трубки, компактно покладеної навколо гема (небілковий компонент, що містить залізо)
Першим білком, третинна структура якого була з'ясована Дж. Кендра на підставі рентгеноструктурного аналізу, виявився миоглобин кашалота. Це порівняно невеликий білок з мовляв. м. 16700, що містить 153 амінокислотних залишку (повністю з'ясована первинна структура), представлений однією поліпептидного ланцюгом. Основна функція міоглобіну - перенесення кисню в м'язах. Поліпептидний ланцюг міо-глобіну представлена у вигляді зігнутої трубки, компактно покладеної навколо гема (небілковий компонент, що містить залізо)
Модель третинної структури молекули міоглобіну (по Дж. Кендра). Латинськими літерами позначені структурні домени, червоним кольором - гем Слайд №17  Якщо поліпептидний ланцюг містить більше 200 амінокислот, то її просторова структура сформовані у вигляді доменів.
Якщо поліпептидний ланцюг містить більше 200 амінокислот, то її просторова структура сформовані у вигляді доменів.
Домен - це компактна глобулярна структурна одиниця всередині поліпептидного ланцюга.
Домени можуть виконувати різні функції і піддаватися складанню (згортання) в незалежні компактні глобулярні структурні одиниці, з'єднані між собою гнучкими ділянками всередині білкової молекули.
Відкрито багато білків (наприклад, імуноглобуліни), що складаються з різних за структурою і функціями доменів, кодованих різними генами.
Домени визначаються на генетичному рівні - екзонами. слайд №18 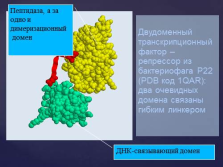 Двудоменний транскрипційні фактор - репрессор з бактеріофага P22 (PDB код 1QAR): два очевидних домену пов'язані гнучким лінкером
Двудоменний транскрипційні фактор - репрессор з бактеріофага P22 (PDB код 1QAR): два очевидних домену пов'язані гнучким лінкером
ДНК-зв'язуючий домен
Пептідаза, а за одне і дімерізаціонний
домен Слайд №19  Під четвертичной структурою мають на увазі спосіб укладання в просторі окремих поліпептидних ланцюгів, що володіють однаковою (або різною) первинної, вторинної або третинної структурою, і формування єдиного в структурному і функціональному відносинах макромолекулярної освіти.
Під четвертичной структурою мають на увазі спосіб укладання в просторі окремих поліпептидних ланцюгів, що володіють однаковою (або різною) первинної, вторинної або третинної структурою, і формування єдиного в структурному і функціональному відносинах макромолекулярної освіти.
Кожна окремо взята поліпептидний ланцюг, що отримала назву протомеров, мономера або субодиниці, найчастіше не володіє біологічною активністю.
Цю здатність білок набуває при певному способі просторового об'єднання входять до його складу протомеров, тобто виникає нова якість, не властиве мономірним білку. Утворену молекулу прийнято називати олігомером. слайд №20  олігомерні білки
олігомерні білки
Олігомерні білки частіше побудовані з парного числа протомеров (від 2 до 4, рідше від 6 до 8) з однаковими або різними молекулярними масами - від декількох тисяч до сотень тисяч.
Зокрема, молекула гемоглобіну складається з двох однакових? - �� двох? -Поліпептідних ланцюгів, тобто являє собою тетрамер. слайд №21 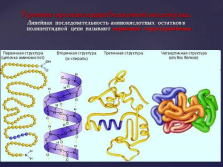 Рівні організації білкової молекули.
Рівні організації білкової молекули.
Лінійна послідовність амінокислотних залишків у поліпептидному ланцюзі називають первинною структурою білка. слайд №22 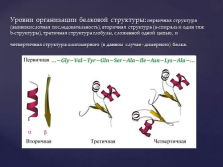 Рівні організації білкової структури: первинна структура (амінокислотна послідовність), вторинна структура (a-спіраль і один тяж b-структури), третинна структура глобули, складеної одним ланцюгом, і четвертинна структура олігомерного (в даному випадку - дімерная) білка. слайд №23
Рівні організації білкової структури: первинна структура (амінокислотна послідовність), вторинна структура (a-спіраль і один тяж b-структури), третинна структура глобули, складеної одним ланцюгом, і четвертинна структура олігомерного (в даному випадку - дімерная) білка. слайд №23  Таким чином, лінійна одномірна структура поліпептидного ланцюга (тобто послідовність амінокислотних залишків, обумовлена кодом білкового синтезу) наділена інформацією іншого типу - конформационной, яка являє собою утворення білкової молекули строго заданої форми з певним просторовим розташуванням окремих її частин. слайд №24
Таким чином, лінійна одномірна структура поліпептидного ланцюга (тобто послідовність амінокислотних залишків, обумовлена кодом білкового синтезу) наділена інформацією іншого типу - конформационной, яка являє собою утворення білкової молекули строго заданої форми з певним просторовим розташуванням окремих її частин. слайд №24  Фолдінг - процес просторової укладки синтезованої поліпептидного ланцюга, формування єдино можливої нативной структури білка. У клітинах відбувається відбір з безлічі стерически можливих станів однієї-єдиної стабільної і біологічно активної конформації, яка визначається, найімовірніше, первинною структурою. У фолдінг беруть участь білки - шаперони. І хоча більшість щойно синтезованих білків можуть згортатися і при відсутності шаперонов, деякого меншості обов'язково потрібно їх присутність. слайд №25
Фолдінг - процес просторової укладки синтезованої поліпептидного ланцюга, формування єдино можливої нативной структури білка. У клітинах відбувається відбір з безлічі стерически можливих станів однієї-єдиної стабільної і біологічно активної конформації, яка визначається, найімовірніше, первинною структурою. У фолдінг беруть участь білки - шаперони. І хоча більшість щойно синтезованих білків можуть згортатися і при відсутності шаперонов, деякого меншості обов'язково потрібно їх присутність. слайд №25  Шаперони. слайд №26
Шаперони. слайд №26 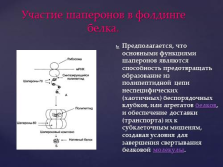 Участь шаперонов в згортання білків.
Участь шаперонов в згортання білків.
Передбачається, що основними функціями шаперонов є здатність запобігати утворенню з поліпептидного ланцюга неспецифічних (хаотичних) безладних клубків, або агрегатів білків, і забезпечення доставки (транспорту) їх до субклітинних мішенях, створюючи умови для завершення згортання білкової молекули. слайд №27 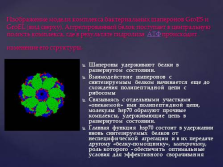 Зображення моделі комплексу бактеріальних шаперонов GroES і GroEL (вид зверху). Агрегованих білок надходить в центральну порожнину комплексу, де в результаті гідролізу АТФ відбувається зміна його структури.
Зображення моделі комплексу бактеріальних шаперонов GroES і GroEL (вид зверху). Агрегованих білок надходить в центральну порожнину комплексу, де в результаті гідролізу АТФ відбувається зміна його структури.
Шаперони утримують білки в розгорнутому стані.
Взаємодія шаперонов з синтезованим білком починається ще до сходження поліпептидного ланцюга з рибосоми
Зв'язуючись з окремими ділянками «опікувався» ними поліпептидного ланцюга, молекули hsp70 утворюють міцні комплекси, які утримують ланцюг в розгорнутому стані.
Головна функція hsp70 полягає в утриманні знову синтезованих білків від неспецифічної агрегації і в їх передачі іншій «білку-помічнику», шапероніну, роль якого - забезпечити оптимальні умови для ефективного згортання
слайд №28  Знову синтезований білок або поліпептид не завжди функціонально активні і вимагають додаткових перетворень, що включають:
Знову синтезований білок або поліпептид не завжди функціонально активні і вимагають додаткових перетворень, що включають:
Фолдінг молекул.
Освіта дисульфідних містків між залишками цистеїну.
Частковий протеоліз.
Приєднання простетичної групи. Збірка протомеров в олігомерного білок.
Модифікацію амінокислотних залишків: фосфорилирование, гідроксилювання і інші реакції.
Пострансляціонная модифікація білків Слайд №29  Деградація білків - Убіквітіновий сигнальний шлях Білок виконує закріплену за ним функцію, а потім, в певний момент, клітині необхідно від нього позбавитися. Останнє обумовлено рядом причин: по - перше, подальша активність білка може нашкодити клітці, по - друге, потрібно синтезувати нові білки, а перевантаження цитоплазми поліпептидами є джерелом апоптоза.Внутріклеточную деградацію білків довгий час вважали неспецифічним випадковим процесом. Справжнім проривом в даній області послужило відкриття убіквітінового сигнального шляху. В рамках цього шляху деградації білка, яка здійснюється великим білковим комплексом - протеосомой, передує приєднання до нього «ланцюжка» молекул невеликого пептиду убіквітину. слайд №30
Деградація білків - Убіквітіновий сигнальний шлях Білок виконує закріплену за ним функцію, а потім, в певний момент, клітині необхідно від нього позбавитися. Останнє обумовлено рядом причин: по - перше, подальша активність білка може нашкодити клітці, по - друге, потрібно синтезувати нові білки, а перевантаження цитоплазми поліпептидами є джерелом апоптоза.Внутріклеточную деградацію білків довгий час вважали неспецифічним випадковим процесом. Справжнім проривом в даній області послужило відкриття убіквітінового сигнального шляху. В рамках цього шляху деградації білка, яка здійснюється великим білковим комплексом - протеосомой, передує приєднання до нього «ланцюжка» молекул невеликого пептиду убіквітину. слайд №30  Поліубіквітіновая ланцюжок навішується в строго певний момент і є сигналом, що свідчить про те, що даний білок підлягає деградації. Амінокислота, по залишку якої убиквитин зв'язується з білками - лізин. Тепер ясно, що процес внутрішньоклітинного протеолізу жорстко регулюється і надзвичайно важливий для безлічі базальних клітинних функцій. Серед субстратів специфічного протеолізу: регулятори клітинного циклу, компоненти різних сигнальних шляхів, а також мутантні білки і білки, пошкоджені посттрансляційних.
Поліубіквітіновая ланцюжок навішується в строго певний момент і є сигналом, що свідчить про те, що даний білок підлягає деградації. Амінокислота, по залишку якої убиквитин зв'язується з білками - лізин. Тепер ясно, що процес внутрішньоклітинного протеолізу жорстко регулюється і надзвичайно важливий для безлічі базальних клітинних функцій. Серед субстратів специфічного протеолізу: регулятори клітинного циклу, компоненти різних сигнальних шляхів, а також мутантні білки і білки, пошкоджені посттрансляційних.
3. Містять тільки?
Зокрема, молекула гемоглобіну складається з двох однакових?
? двох?











