
Просторова організація ДНК
С.В. Разін
Сергій Володимирович Разін, член-кореспондент РАН, доктор біологічних наук,
завідувач лабораторією структурно-функціональної організації хромосом в Інституті біології гена РАН,
професор кафедри молекулярної біології біологічного факультету МГУ им.М.В.Ломоносова.
Ще на початку минулого століття завдяки використанню чисто генетичних методів з'ясувалося, що гени лінійно розташовані на хромосомах. З тих пір більшість дослідників розглядають геном як ланцюг послідовно розташованих генів і міжгенних ділянок, що включають різні регуляторні та інші (здавалося б незначні) послідовності. Такий стереотип мислення відбивається, зокрема, в тому, що відстані між генами або іншими ділянками ДНК зазвичай вказують в тисячах нуклеотидних пар, маючи на увазі відстані уздовж молекули ДНК.
Хоча це цілком коректно, але таке уявлення про лінійність генома містить в собі певні небезпеки. Справа в тому, що в ядрі еукаріотичної клітини геном упакований надзвичайно складно. В результаті послідовності ДНК, в тому числі і гени, віддалені один від одного на десятки або сотні тисяч нуклеотидних пар, а іноді і взагалі розташовані в різних хромосомах, в тривимірному просторі виявляються в безпосередній близькості. Це забезпечує взаємодію білкових комплексів, пов'язаних з віддаленими (якщо вважати уздовж молекули ДНК) регуляторними елементами. Такі взаємодії значно розширюють можливості роботи різних регуляторних систем в геномі еукаріотичної клітини. В останні роки отримано кілька принципово нових спостережень, істотно підвищили інтерес до просторової організації ДНК в ядрі. Ми спробуємо підсумувати сучасні досягнення в цій галузі.
Упаковка ДНК в ядрі
У середній еукаріотичної клітці загальна протяжність геномної ДНК становить близько 2 м, діаметр її ядра всього ~ 10-20 мкм. При цьому сукупність генів, які працюють в даній клітині, повинна бути доступна для РНК-полімерази і транскрипційних факторів, а вся ДНК в клітинах, що діляться повинна реплицироваться.
Сьогодні відомо, що упаковка ДНК в ядрі еукаріотичної клітини здійснюється в кілька етапів (рис.1). Спочатку нитку ДНК укладається в нуклеосоми, при цьому її довжина зменшується в шість-сім разів. Потім нуклеосомна нитка складається в так звану 30 нм фибриллу (соленоїд або зигзагоподібну нитка), що забезпечує додаткову компактизації в 40 разів. Далі фібрила організовується в великі (50 і більше тисяч пар нуклеотидів) петлі, кінці яких закріплюються на білковому скелеті ядра (його часто називають ядерним матриксом). На цьому етапі лінійні розміри ДНК скорочуються в 700 раз [ 1 ]. Існують і такі рівні компактизації ДНК, інформація про яких в даний час дуже мізерна і суперечлива.
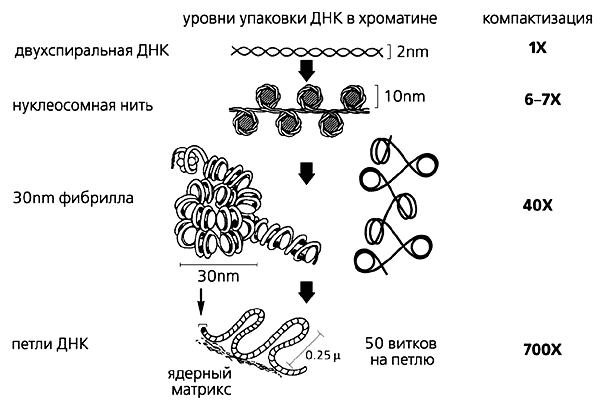
Мал. 1. Рівні упаковки ДНК в ядрі еукаріотичної клітини.
Поки мова йшла лише про упаковку однієї протяжної молекули ДНК. У першому наближенні такий можна вважати ДНК однієї хромосоми. Однак геном еукаріотичної клітини розділений на кілька хромосом. Наприклад, в клітинах улюбленого об'єкта генетиків - плодової мушки дрозофіли - є чотири пари хромосом (в клітинах людини їх 46). Індивідуальні хромосоми можна побачити під мікроскопом тільки під час мітозу. На інших фазах клітинного циклу вони не видно, і ядро клітини представляється щодо гомогенним. Протягом багатьох років молекулярних біологів цікавило питання, займають чи окремі хромосоми обмежені простору всередині ядра або ж при декомпактізаціі хромосом ДНК кожної з них розподіляється по всьому ядру, неминуче перемішуючись з ДНК інших хромосом.
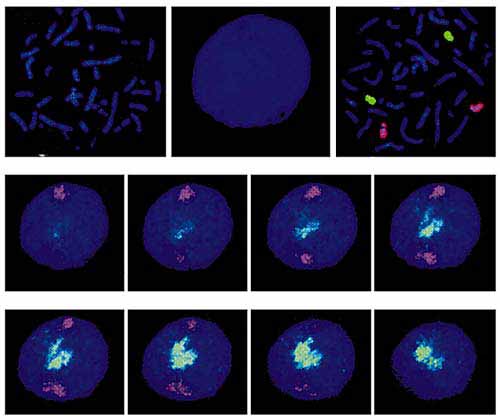
Мал. 2. Забарвлення хромосомних територій.
А - ДНК метафазних хромосом людини.Б - ДНК інтерфазних ядра.В - ДНК метафазних хромосом (синій колір) після гібридизації з хромосом-специфічними пробами, дізнавшись хромосому 18 (червоний колір) і хромосому 19 (зелений колір);показані два гомолога відповідної хромосоми.Г - результати гібридизації ДНК інтерфазних ядер з хромосом-специфічними пробами, дізнавшись хромосоми 18 і 19. Вісім секцій ядра зроблені за допомогою конфокального мікроскопа;синім пофарбована вся ядерна ДНК (як на ріс.б).Близько 10 років тому відповідь на це питання було знайдено. Методи молекулярної гібридизації дозволили фарбувати в інтерфазних ядрі індивідуальні хромосоми (рис.2). Виявилося, що вони, всупереч загальноприйнятій в той час точці зору, займають всередині ядра обмежені неперекривающіеся простору (названі "хромосомними територіями", рис.3) і розташовуються невипадковим чином: хромосоми, багаті генами, локалізуються ближче до центру ядра, а бідні генами - ближче до його периферії [ 2 ]. У підтримці специфічних позицій хромосомних територій важливу роль відіграє ядерний матрикс.
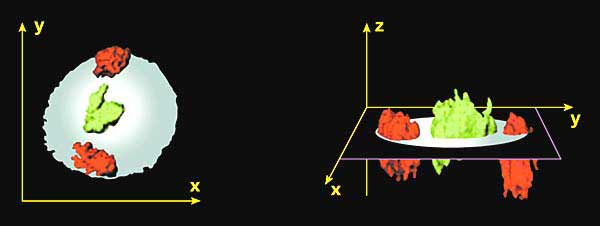
Мал. 3. Двовимірна і тривимірна модель ядра, що показують розташування хромосомних територій [ 2 ].
Взаємодія віддалених регуляторних елементів
Упаковка ДНК в ієрархічні хроматіновие структури принципово важлива для фізичного відстані між регуляторними послідовностями і їх орієнтації в просторі. А ці послідовності завжди служать майданчиками зв'язування регуляторних білків. Уже організація ДНК в нуклеосоми може зробити ці майданчики недоступними для білкових чинників, або орієнтувати їх так, що посаджені на них білкові комплекси в силу чисто стеріческіх причин (наприклад, спрямованості в протилежні сторони) не зможуть взаємодіяти один з одним. А при формуванні фібрил можливості придушення або активації тих чи інших регуляторних систем зростають. Однак розташування нуклеосом на ДНК досить динамічно. Зворотні зміни в їх структурі і ступеня конденсації хроматину (зокрема, перехід від розгорнутої нуклеосомної нитки до 30 нм фібрили і більш компактним гетерохроматинових структурам) складають найбільш вивчену частина епігенетичних механізмів.
Ці механізми ми не будемо обговорювати, а зупинимося на наступному рівні упаковки ДНК в хроматині, а саме на протяжних петлях ДНК (рис.1). Їх можна побачити при електронній мікроскопії метафазних хромосом і інтерфазних ядер, з яких були видалені гістони. Наявність в інтерфазних ядрах топологічно замкнутих петель ДНК продемонстровано і за допомогою біохімічних методів [ 4 ].
Прикріплені до ядерного матриксу петлі ДНК зацікавили фахівців перш за все тому, що за своїми розмірами вони могли б відповідати функціональним одиницям генома. Для перевірки цього припущення було потрібно вивчити специфічність організації ДНК в петлі. По-перше, встановити, чи однаково поділ генома на петлі у всіх клітинах. Якщо це припущення вірне, то одні фрагменти геному завжди повинні знаходитися в підставах петель ДНК, а інші - в самих петлях. По-друге, з'ясувати, чи є якісь особливі послідовності ДНК, відповідальні за "заякоріванню" петель на білковому матриксі ядра.
Метод розрізання генома
За останні 30 років запропоновано декілька методів картування ділянок прикріплення петель ДНК до ядерного матриксу (хромосомному остову). Хоча ці методи розрізняються в деталях, їх можна розділити на дві принципово різняться групи. Перша заснована на виділенні так званої "прилеглої до ядерного матриксу ДНК" (тобто знаходиться в підставах петель) і фракції петель ДНК, яка відщеплюється від ядерного матриксу при обмеженою обробці ядер ферментом нуклеаз (рис.4). Бажаний присутність досліджуваного фрагмента ДНК в прилеглій ядерного матриксу фракції, отриманої після досить інтенсивної нуклеазного обробки, дозволяє говорити про те, що він прикріплений до ядерного матриксу. Друга група методів спрямована на вивчення специфічності послідовностей ДНК, що взаємодіють з ядерним матриксом. В основі всіх методів лежить виборче зв'язування in vitro (тобто в пробірці) фрагментів клонованої ДНК з ізольованим ядерним матриксом. Однак результати, отримані за допомогою двох методичних підходів, виявилися досить суперечливими [ 4 ].
![Однак результати, отримані за допомогою двох методичних підходів, виявилися досить суперечливими [ 4 ]](/wp-content/uploads/2020/01/uk-vivos-voco-sv-razin-quotprostorova-organizacia-dnkquot-4.GIF)
Мал. 4. Схема встановлення позицій генів в петлях ДНК.
А - нуклеоїд, отриманий після екстракції гістонів з ядер, які не оброблених нуклеазами;одна з петель ДНК містить три гени: "a" знаходиться в проксимальній (по відношенню до ядерного матриксу) частини петлі, "b" займає проміжне положення, і "c" - в дистальній частині;
Б - після обробки ядер нуклеазами утворюються приблизно два розриву на петлю; фрагменти ДНК, що знаходяться в підставах петель (всередині пунктирного кола), прикріплені до ядерного матриксу. Після диференціального центрифугування фрагменти поділяються, і в прикріпленою до ядерного матриксу ДНК залишаються гени "a" і "b";
В - після додаткової обробки нуклеазами залишається тільки ген "а".
Ми звернули увагу, що всі методи спрямовані на ідентифікацію та характеристику фрагментів ДНК, локалізованих в підставах петель. В основі нашого принципово нового підходу лежить розрізання всього генома на петлі і подальша їх характеристика. На перший погляд, розділити геном на індивідуальні петлі надзвичайно важко. Тут дуже важливо, за допомогою якого інструменту робити розриви в підставах петель ДНК. На щастя, таким інструментом нас забезпечила сама природа. Дослідження показали, що один з головних компонентів ядерного матриксу - фермент ДНК-топоізомераза II, який регулює топологію ДНК. Цей фермент вносить двунітевие розриви в ДНК, які після зняття топологічних напруг або поділу катенанів зашиваються (лігуються) тим же ферментом. Протягом всієї реакції фермент, що складається з двох субодиниць, залишається пов'язаним з ДНК.
Існує цілий ряд інгібіторів ДНК-топоізомерази II (в нашому випадку VM-26), які зупиняють реакцію на стадії проміжного комплексу фермент-ДНК. (Цікаво, що більшість з них використовуються в якості протипухлинних агентів.) При цьому кожна з субодиниць ферменту залишається ковалентно пов'язаної з 5 ў-кінців розірваного ланцюга ДНК. Якщо такі блоковані комплекси обробити денатуруючих агентом, після чого зруйнувати фермент, то вийде препарат ДНК, розрізаний на фрагменти в місцях контакту ДНК з ферментом (рис.5). Якби топоізомераза II перебувала тільки в ядерному матриксі, то проста обробка живих клітин її інгібіторами розрізала б весь геном по ділянках прикріплення ДНК до ядерного матриксу. Однак завдання ускладнюється тим, що цей фермент в розчинній формі присутній в нуклеоплазмі і може вносити розриви в будь-якому місці (якщо виявиться поруч з ДНК в момент обробки клітин інгібітором). Найбільш ймовірними точками розривів будуть вільні від нуклеосом ділянки, найбільш чутливі до ДНК-нуклеаз (ДНКаза I). Щоб виключити можливість розривів поза цікавлять нас ділянок прикріплення ДНК до ядерного матриксу, ми екстрагували розчинний фермент, а заодно і гістони, обробляючи ядра 2M NaCl (рис.4). Отримані так звані нуклеоїди обробляли інгібіторами ДНК-топоізомерази II [ 5 ]. Так нам вдалося розрізати весь геном на окремі петлі і їх олігомери.
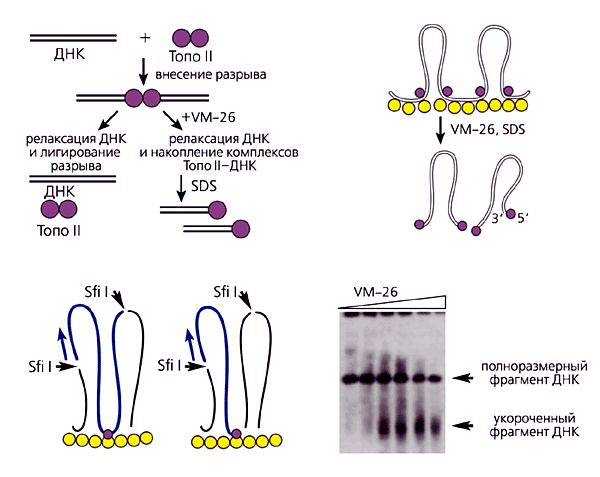
Мал.5. Схема методу картування петель ДНК:А - реакція, що каталізується ДНК-топоізомеразою II, і механізм розрізання ДНК при інгібуванні зшивається активності ферменту VM-26 і іншими "топоізомеразнимі отрутами";
Б - розрізання геномної ДНК на індивідуальні петлі. Після видалення гістонів розгорнуті петлі ДНК все ще прикріплені до ядерного матриксу (жовті гуртки), який містить ДНК-топоізомеразу II (фіолетові кружки). Нуклеоїди инкубируют в середовищі з VM-26, після чого лизируют додецилсульфатом натрію (SDS). У місцях прикріплення петель до ядерного матриксу топоізомераза II розрізає ДНК;
В - в петлях ДНК, оброблених рестриктазой Sfi I, з'являються розриви. Фрагмент Sfi I-Sfi I (показаний синім кольором) можна ідентифікувати за допомогою гібридизації з пробою, комплементарної одному з кінців повнорозмірного рестріктного фрагмента (синя стрілка). Справа ту саму ділянку генома після додаткового розриву, викликаного топоізомеразою II (фіолетовий гурток). Розмір укороченого фрагмента дорівнює відстані від ділянки розщеплення ДНК-рестриктазой Sfi I до ділянки розщеплення її топоізомеразою II;
Г - типова картина результату електрофорезу. На всіх доріжках видно повнорозмірний Sfi I-Sfi I фрагмент ДНК. В доріжках, що містять ДНК з нуклеоидов, оброблених високими концентраціями VM-26, з'являється додатковий (Sfi I-Тopo II) фрагмент, який свідчить, що всередині досліджуваного фрагмента ДНК знаходиться ділянка прикріплення до ядерного матриксу.
Що робити далі? Як встановити позиції решт петель на фізичній карті генома? Нагадаємо, що на такій карті показані реальні відстані уздовж молекули ДНК між тими чи іншими маркерами. Фізичні карти різних геномів почали створювати задовго до розшифровки геномів людини і ряду інших організмів. В якості маркерів при створенні таких карт зазвичай використовуються ділянки розщеплення ДНК ферментами рестріктазамі. Встановити позиції ділянки прикріплення петлі ДНК до ядерного матриксу на фізичній карті - значить визначити відстань від місця прикріплення до місця розщеплення ДНК тій чи іншій рестриктазой. Для цього можна скористатися методом непрямого мічення решт фрагментів ДНК [ 6 ], Запропонованим близько 30 років тому для картування позицій ділянок гіперчутливості до ДНКаза I.
Принцип цього методу полягає в тому, що після внесення в ДНК розривів тим чи іншим агентом (в нашому випадку - ДНК-топоізомеразою II ядерного матриксу) препарат додатково розрізають обраної рестриктазой. Після поділу фрагментів за допомогою електрофорезу і перенесення їх на нітроцелюлозний фільтр проводять гібридизацію з пробою, комплементарної кінця вирізаного фрагмента, всередині якого може знаходитися додатковий розрив. Якщо такого розриву немає, то після гібридизації вийде повнорозмірний фрагмент. Але якщо всередині цього фрагмента ДНК була розрізана топоізомеразою II ядерного матриксу або іншим ферментом, фрагмент буде коротшим, і довжина його дорівнює відстані від ділянки розщеплення ДНК рестриктазой до ділянки розщеплення ДНК досліджуваних агентом (рис.5). При роботі з петлями, вирізаними ДНК-топоізомеразою II, основна складність полягає в необхідності розділити за розміром дуже довгі фрагменти ДНК. Цю проблему можна вирішити, використовуючи електрофорез в пульсуючому полі, який дозволяє розділити фрагменти ДНК з розмірами від декількох тисяч до декількох мільйонів нуклеотидних пар [ 7 ].
Карта організації в петлі-домени гена дистрофина людини
Ми з успіхом використовували вищеописаний метод для картування меж петель в ряді областей геному людини і дрозофіли. Після цього була поставлена масштабна задача - побудувати карту організації в петлі-домени найдовшого з відомих генів - гена дистрофина людини. У цьому гені, розташованому на Х-хромосомі, близько 2500 тис. Нуклеотидних пар, а розмір його мРНК складає всього 14 тис. Нуклеотидних пар. Інакше кажучи, більше 99% від загальної протяжності гена займають некодуючі послідовності (інтрони). У гені дистрофина часто відбуваються різні перебудови, деякі з яких призводять до тяжких спадкових захворювань - м'язовим дистрофій [ 8 ].
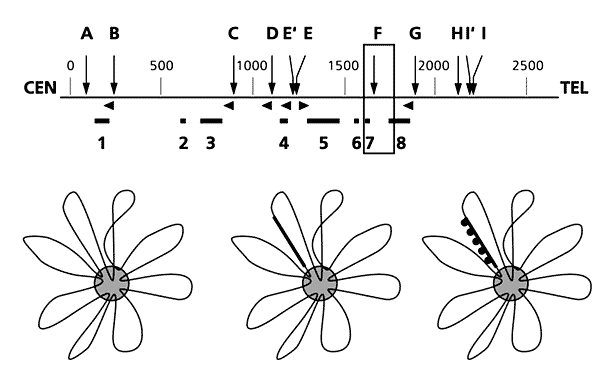
Мал.6. Карта організації в петлі гена дистрофина людини.Вгорі - схема розташування ділянок і областей прикріплення ДНК до ядерного матриксу (горизонтальні лінії, позначені номерами 1-8) в межах гена дистрофина.Вертикальні стрілки (латинські літери AI) вказують на розташування ділянок розщеплення;горизонтальні - на позиції гібрідізаціонних проб.
Внизу - схема візуалізації унікальних фрагментів ДНК на препаратах нуклеоидов. Після екстракції з ядер гістонів петлі ДНК розправляються і утворюють корону навколо ядерного матриксу (a). Препарати гибрідизуючою з пробами, що містять біотин (жирна смуга на схемі б). Таку пробу можна побачити після забарвлення антитілами, пов'язаними з флуоресцентним барвником (чорні кружечки на схемі в).
Карту розщеплення гена дистрофина рестриктазой SfiI побудував ще до визначення повної нуклеотідної послідовності генома людини. Ми картірован позіції ділянок Прикрепление ДНК до ядерного матриксу относительно точок розщеплення ДНК цієї рестриктазой и з'ясували, что в гені дистрофина є щонайменш дев'ять петель, розділеніх вісьмома зонами Прикрепление [ 9 ]. У Деяк випадка протяжність ділянок ДНК, что розділяють две сусідні петлі, порівнянна з довжина самих петель (рис.6, а). Це принципова нове спостереження дозволило розглядаті зони Прикрепление ДНК до ядерного матриксу як особливо часть генома. Цікаво, что именно тут знаходяться віявлені Ранее в гені дистрофина гарячі точки рекомбінації [ 9 ]. Виявлено закономірність виявило справедливою и для ряду других Вивчення генів. Ще одне цікаве спостереження (також підтверджене на інших експериментальних моделях) полягає в тому, що в зонах прикріплення петель розташовуються ділянки, з яких починається реплікація ДНК. Це підтверджує сформульоване нами ще 20 років тому положення про найважливіший принцип організації еукаріотичної хромосоми - її побудові з структурно-функціональних доменів, відповідних репликационная одиницям [ 10 ].
Петлі ДНК під мікроскопом
Експериментальний підхід, використаний нами для картировании петель ДНК, заснований на ряді логічних передумов, що випливають з радіально-петлевий моделі будови хромосоми. До недавнього часу не було прямих доказів того, що петлі ДНК, картірованниє за допомогою різних методів, саме ті, які можна бачити на цитологічних препаратах. Серед безлічі переплетених петель ДНК, які спостерігаються під електронним мікроскопом, практично неможливо ідентифікувати петлю як фрагмент генома, що цікавить дослідника. Однак це можливо при аналізі петель з більш низьким дозволом.

Мал.7. Мікрофотографії результатів гібридизації in situ з препаратами ядерних гало (a) з фрагментом людського генома - петлі ДНК, картірован в гені дистрофина.Ця петля обмежена областями прикріплення до ядерного матриксу 7 і 8. Гібридизація без конкурентної ДНК (б) і в присутності надлишку немічених фракції повторюванихпослідовностей людської ДНК (в).
Якщо подивитися на екстраговані 2M NaCl ядра в флуоресцентного мікроскопі (після фарбування ДНК тих чи інших флуоресцентним барвником), то можна бачити корону петель ДНК у вигляді хмари, що оточує більш яскраво забарвлену центральну зону (ядерний матрикс) (рис.7, а й схеми на рис.6). Такі препарати називають ядерними гало (nuclear halos), на яких індивідуальні петлі невиразні. Щоб побачити їх, треба скористатися методом гібридизації in situ (в даному випадку препаратів іммобілізованих на склі ядерних гало) з цікавлять фрагментом геному. Проба повинна містити аналоги нуклеотидів (наприклад біотінілірованние уридин), які після гібридизації фарбуються флуоресцентними барвниками, наприклад червоним або зеленим. Це дозволяє одночасно аналізувати розподіл ДНК, яку найпростіше забарвити DAPI (4 ', 6-діамідинів-2-феніліндолом) в бузковий колір, і розподіл проби після гібридизації, пофарбованої в червоний або зелений колір.
В ході реалізації програми з секвенування генома людини в різних лабораторіях клонували тисячі протяжних (100-300 тис. Нуклеотидних пар) фрагментів ДНК людини. Більшість клонів систематизували відповідно позиціями клонованих фрагментів ДНК в геномі людини. Існує цілий ряд наукових центрів, в яких можна придбати цікавить клон. Ми взяли клонований фрагмент людської ДНК, що представляє картірован нами в гені дистрофина петлю ДНК, обмежену ділянками прикріплення 7 і 8 (див. Рис.6). Після гібридизації цього фрагмента з препаратами ядерних гало виявляється безліч сигналів, розподілених по всьому полю (рис.7, б). Це пов'язано з тим, що в ДНК вищих еукаріот, в тому числі і людини, присутня безліч повторюваних послідовностей, розподілених по всьому геному.
Наявні в нашій пробі повтори гибрідизуючою з усіма комплементарними послідовностями. Зрозуміло, що результати такого експерименту не піддаються інтерпретації. На щастя, сигнали від гібридизації повторюванихпослідовностей можна придушити. Для цього ми провели гібридизацію в присутності надлишку немічених фракції повторюваних послідовностей ДНК людини і побачили петлі ДНК, прикріплені до ядерного матриксу (рис.7, в). Всі вони мали однаковий розмір (в межах похибки вимірювань), відповідний довжині фрагмента ДНК, картірован в експериментах з вирізання петель ДНК-топоізомеразою II ядерного матриксу [ 9 ].
Значення цього результату виходить за рамки простого підтвердження правильності побудованої нами карти доменної організації гена дистрофина. Вперше в світі ми показали, що біохімічний метод, заснований на радіальної моделі будови хромосоми, дійсно дозволяє картировать петлі ДНК, які спостерігаються на цитологічних препаратах. Це підтверджує і радіальну модель будови хромосоми, на підставі якої розроблено наш метод вирізування петель. Далі, можливість спостерігати однакові петлі ДНК при аналізі ряду препаратів ядерних гало підтверджує той факт, що ДНК організована в петлі статично, тобто у всіх клітинах до ядерного матриксу прикріплені одні і ті ж фрагменти ДНК, а ділянки між ними утворюють петлі. Ми поставили експеримент на активно діляться клітинах. Коль скоро у всіх клітинах виявлені однакові петлі ДНК, можна стверджувати, що специфічна організація ДНК в петлі, розділені зонами прикріплення, зберігається в ряді клітинних поділів. Ця обставина надзвичайно важливо, оскільки дозволяє розглядати таку організацію ДНК як один з епігенетичних механізмів. Дійсно, при утворенні петель можуть фіксуватися позиції різних регуляторних елементів і їх мішеней, сприяючи їх взаємодії або, навпаки, виключаючи його.
Петлі ДНК, хромосомні перебудови і еволюція геному
Як ми вже говорили, гарячі точки рекомбінації гена дистрофина знаходяться в сегментах, прикріплених до ядерного матриксу. Додаткові дослідження показали, що до ядерного матриксу прикріплені і гарячі точки рекомбінації, присутні в ряді інших генів, зокрема тих, рекомбінації яких асоційовані з розвитком лейкозів [ 11 ]. Важко повірити, щоб це було просто випадковим збігом. Швидше за все, саме постійний контакт ДНК з топоізомеразою служить причиною виникнення "гарячих точок" хромосомних перебудов. Топоізомераза II може прямо брати участь в незаконній рекомбінації. Ще більш імовірно, що внесені нею двунітевие розриви в ДНК при певних умовах можуть стимулювати неточне відновлення цих пошкоджень.
Відомо, що репарація двунітевих розривів в ДНК вищих еукаріот нерідко призводить до різних рекомбінаційним подій. На можливу роль топоізомерази II як індуктора хромосомних перебудов вказують численні дані про те, що використання інгібіторів цього ферменту в хіміотерапії пухлин нерідко викликає вторинні лейкози [ 12 ]. Клітини цих лейкозів характеризуються різними великомасштабними хромосомними змінами, найбільш частими в ділянках розщеплення ДНК-топоізомеразою II [ 13 ]. Важливо відзначити, що ділянки прикріплення петель на молекулі ДНК розташовуються досить далеко один від одного, але всередині ядра вони можуть виявитися в безпосередній близькості. Незаконна рекомбінація між такими ділянками буде приводити до втрати або переміщенню протяжних ділянок геному, що, в свою чергу, може бути важливим фактором еволюції геному [ 14 ].
література
1. Getzenberg RH, Pienta KJ, Ward WS, Coffey DS // Journal of Cellular Biochemistry. 1991. V.47. P.289-299.
2. Cremer T., Kurz A., Zirbel R., Dietzel S. et al. // Cold Spring Harb. Symp. Quant. Biol. 1993. V.58. P.777-792.
3. Peterson CL, Laniel MA // Curr. Biol. 2004. V.14. P.R546-551.
4. Razin S., Gromova II, Iarovaia OV // International Review of Cytology. 1995. V.162B. P.405-448.
5. Razin SV, Hancock R., Iarovaia O., Westergaard O. et al. // Cold Spring Harbor Symp. Quant. Biol. 1993. V.58. P.25-35.
6. Nedospasov SA, Georgiev GP // Biochem. Biophys. Res. Commun. 1980. V.92. P.532-539.
7. Schwartz DC, Cantor CR // Cell. 1984. V.37. P.67-75.
8. Hoffman EP, Schwartz L. // Mol. Aspects Med. 1991. V.12. P.175-194.
9. Iarovaia OV, Bystritskiy A., Ravcheev D., Hancock R. et al. // Nucl. Acids. Res. 2004. V.32. P.2079-2086.
10. Razin SV, Kekelidze MG, Lukanidin EM, Scherrer K. et al. // Nucl. Acids Res. 1986. V.14. P.8189-8207.
11. Iarovaia OV, Shkumatov P., Razin SV // J. Cell. Sci. 2004. V.117. P.4583-4590.
12. Super HJ, McCabe NR, Thirman MJ, Larson RA et al. // Blood. 1993. V.82. P.3705-3711.
13. Zhang Y., Strissel P., Strick R., Chen J. et al. // Proc. Natl. Acad. Sci. USA. 2002. V.99. P.3070-3075.
14. Razin SV // Crit. Rev. Eukar. Gene Exp. 1999. V.9. P.279-283.
Що робити далі?
Як встановити позиції решт петель на фізичній карті генома?











