
М и шечно сокращ е ня, вкорочення м'яза, в результаті якого вона проводить механічну роботу. М. с. забезпечує здатність тварин і людини до довільних рухів. Найбільш важлива складова частина м'язової тканини - білки (16,5-20,9%), в тому числі контрактильні, що зумовлюють здатність м'яза до скорочення. Значний інтерес представляють механоактівние миофибриллярних білки, вивчення яких було розпочато В. Кюне (1864). Важливі дані, що характеризують фізико-хімічні та біохімічні властивості механоактівних м'язових білків, були отримані А. Я. Данилевським (1881-88). У 1-ій половині 20 ст. В. А. Енгельгардт і М. Н. Любімова (1939) встановили, що основною контрактільний білок м'язів - міозин - володіє аденозінтріфосфатазную активністю, а А. Сент-Дьyoрдьі і Ф. Б. Штрауб показали (1942-43), що входить до складу міофібрил білок складається в основному з 2 компонентів - міозину і актину . Взаємодія цих фібрилярних білків і лежить в основі феномена скорочення всіляких контрактільних органел та органів руху (див. м'язи ). Періодичні зміна фізичного стану м'язових білків, що обумовлює можливість поперемінного скорочення і розслаблення м'язів і виконання ними механічної роботи, очевидно, пов'язано з певними дають енергію біохімічними процесами. Енгельгардтом і Любимова (1939-42) було виявлено, що особливим чином приготовані з міозину нитки при взаємодії з розчином АТФ різко змінюють свої механічні властивості (еластичність і розтяжність). Одночасно відбувається розщеплення АТФ з утворенням АДФ і неорганічного фосфату. Це відкриття заклало фундамент для нового напряму в біохімії - механохімії М. с. Надалі Сент-Дьyoрдьі і Штрауб показали, що істинним скорочувальним білком є не міозин, а його комплекс з актином - актоміозін . Скороченню при взаємодії з АТФ піддаються як вимочені у воді або 50% -ному гліцерині м'язові волокна, так і нитки, приготовані з актомиозинового гелів (синерезис гелю). Ці досліди підтверджують, що енергія, необхідна для скорочення м'язів, звільняється в результаті взаємодії актомиозина з АТФ з розщепленням останнього на АДФ і H3PO4. При цьому звільняється велика кількість енергії (8-10 ккал, або 33,5-41,9 кДж, на 1 моль АТФ). Однак справжній механізм цієї реакції залишається все ще незрозумілим. Вважають, що термінальна фосфатная угруповання АТФ при взаємодії з актомиозином переноситься на міозин без проміжного освіти тепла з утворенням багатою енергією фосфорильованій форми актомиозина, здатної до скорочення. Молекулярна маса міозину, певна методом ультрацентрифугирования, близька до 500 тис. Молекула міозину може бути розщеплена без розриву ковалентних зв'язків на більш дрібні субодиниці (рис. 1): 2 «важкі» поліпептидні ланцюжки з молекулярною масою понад 210 000 і 2 (по ін. даними, 3) коротких ( «легких») поліпептиду з молекулярною масою близько 20 000 кожен. За даними електронної мікроскопії, молекула міозину складається ніби з 2 частин - потовщеною «головки» і довгого «хвоста». Загальна довжина молекули - близько 1600 А. Велике число відповідно розташованих в просторі макромолекул міозину утворює в поперечнополосатого волокні товсті (міозіновие) нитки. В освіті поперечних містків між товстими (міозіновимі) і тонкими (актиновими) нитками безпосередньо беруть участь, по-видимому, «головки» миозинових молекул. Молекулярна маса мономера актину близька до 46 000 (раніше приймалася близько 70 000). Встановлено і його первинна структура: число, природа і послідовність включення в поліпептидний ланцюг амінокислотних залишків. Молекули фибриллярного актину (Ф-актину) утворені 2 спіральними ланцюжками, що складаються з безлічі намистин - молекул глобулярного актину (мономера актину, або Г-актину). У саркомер поперечнополосатого волокна нитки Ф-актину просторово відмежовані від ниток міозину. Взаємодія систем двох типів ниток здійснюється за рахунок енергії, що звільняється при розщепленні АТФ в присутності іонів Ca2 + (рис. 2). Т. к. При роботі м'язів АТФ постійно споживається, для тривалого здійснення двухфазной м'язової діяльності необхідно безперервне відновлення АТФ - її ресинтез. Ресинтез АТФ з АДФ і H3PO4 містить ряд дають енергію перетворень. Найважливіші з них: 1) перенесення фосфатної групи з фосфокреатину (Крф) на АДФ. Ця реакція забезпечує швидкий, що відбувається вже під час скорочення м'язи, ресинтез АТФ за рахунок споживання креатинфосфату; 2) глікогеноліз, або гліколіз (розщеплення глікогену або глюкози з утворенням молочної кислоти); 3) тканинне дихання (утворення АТФ в мітохондріях м'язових волокон за рахунок енергії окислення головним чином вуглеводів, жирних кислот і ненасичених фосфоліпідів). Деяка кількість АТФ може утворюватися також в результаті міокіназной реакції з АДФ: 2 АДФ> <АМФ + АТФ. Фосфорилювання креатину за рахунок АТФ з утворенням Крф здійснюється в процесі гліколізу і тканинного дихання. Ре-синтез Крф і глікогену відбувається головним чином у фазі відпочинку після розслаблення м'язи. Скелетний м'яз, що знаходиться в анаеробних умовах або в умовах кисневого голодування ( гіпоксії ), Здатна до виконання деякої кількості роботи. Однак стомлення в цих випадках настає значно раніше, ніж в присутності кисню, і супроводжується накопиченням в м'язі молочної кислоти.
А. В. Палладіним, Д. Л. Фердманом, Н. Н. Яковлєвим та ін. Отримані дані про біохімічної суті тренування м'язів. С. Є. Северіним продемонстрована здатність дипептидов (карнозина, анзерина) відновлювати працездатність стомлених м'язів і впливати на передачу нервових імпульсів з нерва на м'яз.
Після скорочення, викликаного роздратуванням з нерва або електричним струмом, м'яз незабаром переходить в розслаблений стан, хоча зміст АТФ в м'язових волокнах майже не змінюється. Встановлено, що міофібрили мають здатність взаємодіяти з АТФ і скорочуватися в її присутності лише при наявності в середовищі іонів Ca2 +. Найбільша скорочувальна активність спостерігається при концентрації Ca2 + близько 10-6-10-5 моль. При зниженні вмісту Ca2 + до 10-7 моль або нижче м'язові волокна втрачають здатність до вкорочення і розвитку напруги (що тягне сили) в присутності АТФ. За сучасними уявленнями, в яка покоїться м'язі концентрація іонів Ca2 + підтримується нижче цієї граничної величини внаслідок їх зв'язування структурами (трубочками і бульбашками) саркоплазматической мережі. Зв'язування - це не проста адсорбція, а активний фізіологічний процес, здійснюваний за рахунок енергії, що звільняється при розщепленні АТФ в присутності іонів Mg. Цей механізм отримав назву Са-насоса (за аналогією з «Натрієвих насосом» ). Т. о., Перебування живий м'язи (при наявності в ній достатньої кількості АТФ) в розслабленому стані - результат зниження під дією Са-насоса концентрації іонів Ca2 + в середовищі, що оточує міофібрили, нижче межі, при якому ще можливо прояв АТФ-азной активності і скоротливості актомиозинового структур волокна. Скорочення волокна при подразненні з нерва (або електричним струмом) - результат раптового зміни проникності і, як наслідок, виходу з цистерн і трубочок саркоплазматической мережі і так званої Т-системи іонів Ca2 + в межфібріллярное простір. Поперечні трубочки Т-системи, розташовані на рівні Z-дисків і містять Ca2 +, повідомляються з поверхневою мембраною волокна; тому хвиля деполяризації швидко поширюється по системі трубочок і досягає глибоко розташованих ділянок волокна. Після затухання нервового імпульсу в результаті дії Са-насоса концентрація Ca2 + в межфібріллярних просторі швидко знижується до порогової величини і м'яз переходить в початковий розслаблений стан, поки новий імпульс не викличе повторення всього циклу. Втрату актомиозином здатності розщеплювати АТФ і скорочуватися при зниженні концентрації іонів Ca2 + нижче 10-7 моль пов'язують з присутністю в контрактильной системі особливого білка - тропоніну. При його відсутності актоміозін реагує in vitro (у пробірці) з АТФ практично і в відсутності Ca2 +. У фізіологічних умовах (in vivo) тропонин - постійний компонент контрактильной системи м'язового волокна (тропонин-тропоміозинового комплекс).
Двофазний механізм М. с. не вичерпується викладеними уявленнями. У деяких комах (жуки, бджоли, мухи, комарі та ін.) Частота скорочень м'язів крил багато вище частоти надходять нервових імпульсів. Ці м'язи підпорядкована нейрогенному, а миогенной ритму. Вони можуть здійснювати коливання (осциллировать) кілька сот разів в 1 сек. Осциляція цих м'язів не пов'язана зі зміною концентрації Ca2 + в саркоплазме м'язових волокон. Можливість автоматичної двофазної діяльності клітинних органел руху в присутності АТФ можна спостерігати на клітинних моделях - сперматозоїдах, миготливого епітелію, ундулірующіх мембранах трипаносом і ін. Осциляції органел руху відбувається зі звичайною для даного виду клітин швидкістю при постійній концентрації Ca і триває до тих пір, поки в розчині зберігається відомий надлишок АТФ. Механізм такої осциляції органел руху, а також міофібрил, мабуть, може бути зрозумілий лише виходячи з існування взаємозв'язку між ферментативної активністю (здатністю розщеплювати АТФ) і станом (конформацией) макромолекул скорочувального субстрату.
Літ. см. при ст. м'язи .
І. І. Іванов.
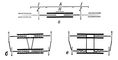
Мал. 2. Гіпотетичні схеми м'язового скорочення: а - конфігурація товстих і тонких ниток в розслабленому м'язі; зміна положення ниток при скороченні: б - модель ковзання (по Х. Хакслі); в - модель скручування (по Подільському).
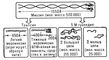
Мал. 1. Структура молекули міозину. Молекула складається з 2 великих і 2 малих (коротких) ланцюгів. Великі ланцюга утворюють довгий «хвіст» молекули; її «головка» складається з кінців великих ланцюгів і 2 малих ланцюгів. (Молекулярна маса окремих ланцюгів визначалася методом ультрацентрифугирования після дезагрегации молекули міозину 5 М солянокислим гуанідином, а також трипсином.)











